My second week in the Maslakova lab has been extremely interesting and an incredible learning experience. This week has been largely focused on learning the art of microinjections - using a fine glass needle to insert a solution of your choice into a living cell. A needle sharp enough and fine enough to pierce a cell without killing it can be created by pulling borosilicate glass capillary tubes with a special "needle pulling" machine. Microinjections are accomplished by carefully maneuvering the needle into a cell - usually egg or early stage embryo - and injecting an extremely small volume of liquid with a pulse of compressed air. The needle is maneuvered about using a micromanipulator - a joystick essentially - and injections are controlled with a foot-pedal, which makes the whole experience feel very much like a video game. Below left, Ph.D. student Laurel Hiebert demonstrates the proper technique.
 |
| The Microinjection Apparatus |
 |
| Fluorescein Labeled Larva. Injection and imaging by Zac Swider |
I initially began doing microinjections with fluorescein - a green fluorescent molecule that is cheap, easy to visualize, and relatively nontoxic. In its own right, fluorescein doesn't do anything interesting (other than make things glow) but if one were to inject a solution of RNA, for example, fluorescein can be co-injected to conveniently mark which cells were injected. Above right is a three day old pilidium larva that I injected with fluorescein as a single cell zygote (a fertilized egg), such that every subsequent cell in the resulting embryo is also labeled with fluorescein. Darker areas indicate parts of the larval body that have undergone more cell division - and thus diluted the dye more so than the brighter areas.
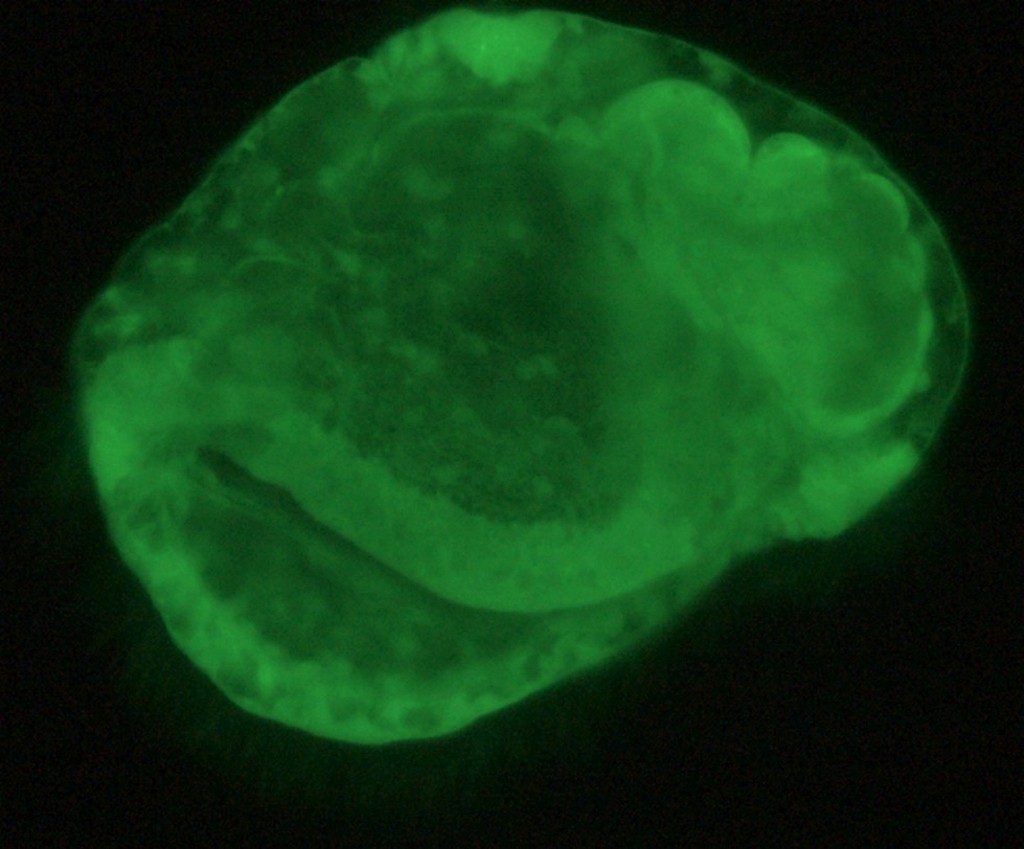
Once I became competent at injecting cells without killing them, we attempted injecting eggs with a solution of mRNA coding for a centrin protein attached to several green fluorescent proteins. Centrin is a component of the centrosome - a crucial bit of intracellular machinery responsible for organizing microtubules within the cell. Microtubules are an essential component of the cell cytoskeleton (the structural scaffolding present within each cell) and so every subsequent cell in the resulting embryo should have a bright green fluorescent component marking it. The cilia that move the larval body around in the water are also largely composed of microtubules and centrin is a component of the basal bodies that generate these cilia. Thus, this labeling technique can be used to differentiate between different cell types that either do not express cilia, express just a few cilia, or express a large amount of cilia. Below is a confocal microscope composition of a three day old nemertean larva expressing centrin-GFP, the above mentioned fluorescent protein, as a result of a microinjection. Confocal microscopy essentially takes a scan of a specimen, from top to bottom, and compiles an image from this scan, eliminating any out of focus light and generating an image far superior to the above "Fluorescein Labeled Larva", which was taken with a compound microscope.
 |
| Centrin-GFP labeled Pilidium Larva. Injection by Zac Swider, imaging by George von Dassow |
The bright clusters of spots in the "armpits" of the above larva are cells with abnormally large "globs" of centrin within them. This phenomenon has been observed before using antibody staining on fixed specimens but never using live imaging. It is known that the cells in the "armpits" (they are technically called the axillary cells) undergo a tremendous amount of division to produce the bulk of the larval body, however, the significance of having such large accumulations of centrin within these cells is completely unknown to science.
Although the process of making things glow in the dark is great fun and it makes for a fantastic picture, my actual research project involves another matter altogether. The Maslakova Lab has a goal of making Micrura alaskensis a model species for nemertean development, and for the next six weeks I will be attempting to characterize the affects of four specific genes on the development of this species. As it turns out, oftentimes the best way to see how something works is to break it and see what goes wrong as a result. This is essentially the approach that I will be taking, and I will be performing morpholino injections to facilitate the "breaking" of these genes. Morpholinos are synthetic (man-made) mRNA analogues that can be made (or in our case, purchased) with a very specific sequence of base pairs that allow them to target and deactivate only the mRNA products of one gene and leave everything else in the cell alone.
 |
| A Cartoon Diagram of a Morpholino Strand Binding to a mRNA Strand (image from Wikipedia.com) |
Essentially - morpholinos block the future production of the protein of your choice. The downside of this of course is that there may already be a significant amount of this protein inside of the cell when you inject the morpholino and it may take some time before the effects of the morpholino become evident.
A few days ago we attempted injections of a morpholino designed to block to the production of the "Mitotic Kinesin-Like Protein-1" (MKLP1) - a motor protein within the cell. Motor proteins generally function to move large objects around within the cell by attaching to them and carrying them to the target location by "walking" along microtubules or actin filaments (another structural component of the cell). MKLP1 also walks along microtubules, but its main function within the cell is to identify where the midline of the cell is - so that the cell can successfully divide into two equal sized daughter cells. When a shortage of this protein occurs, the cells have no hope of dividing further and simply grow larger instead. We observed this expected morphology in M. alaskensis after about 15 hours of development - indicating that M. alaskensis is likely to be responsive to future morpholino treatments as well.
Although this is exciting in itself, in the future we hope to be able to use other fluorescent markers to see exactly what is occurring within these cells as they experience a shortage of MKLP1. The significance of this is as follows: although it is known that MKLP1 is crucial for the ability of cell to identify its midbody, the specifics of what this protein actually does (and the other players that this protein interacts with) are largely still being elucidated by the scientific community. If M. alaskensis proves to be a cooperative research specimen, it could be an invaluable tool in unraveling the mystery of exactly how a cell performs the single most complicated things that it will ever do: divide and become two cells.
 |
| Vials of morpholino solutions for each of the below mentioned genes |
Seen above, morpholino vials for each of the above mentioned "genes of interest". In addition to MKLP1, I will soon be addressing the effects of beta-Catenin and IFT88, more specifics on these to come soon!
Nice to hear the Morpholinos are working in your nemertean - I'm looking forward to the paper!
ReplyDelete- Jon at Gene Tools